◆第3章 複数の色変わり遺伝◆
3-1 はじめに
これまでの解説では,1つの色変わり遺伝についてのみ考察してきました。パイドならパイドだけ,ルチノーならルチノーだけを考察してきたわけです。今回からはこの考察を拡張し,2つの色変わり遺伝子に注目して遺伝の仕組みを考えていくことにしましょう。
既にご承知のように,遺伝の舞台は染色体上です。常染色体上で考えるべき遺伝と性染色体上で考えるべき遺伝とがありました。今,2つの色変わり遺伝子に注目するのですから,我々は次の3つのパターンを考察しなければならないことに気づくでしょう。
1) 常染色体上の遺伝と常染色体上の遺伝の組合せ
2) 常染色体上の遺伝と性染色体上の遺伝の組合せ
3) 性染色体上の遺伝と性染色体上の遺伝の組合せ
1)と2)は,遺伝の仕組みが非常によく似ています。これに対して,3)の場合には,遺伝の仕組みがわずかながら複雑になってくる場合があります。本章では,まず始めに1)と2)について考えていくことにし,最後に3)について少し詳しく考察することにしましょう。3-2 減数分裂の仕組み
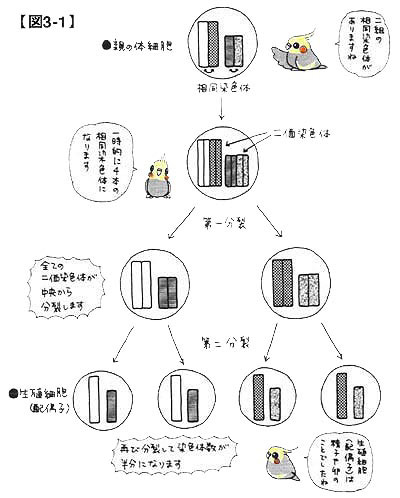
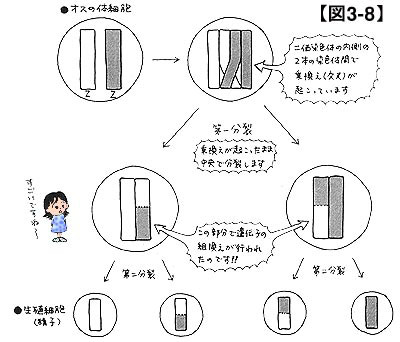
図3-1をご覧ください。これは減数分裂の仕組みを専門用語を用いて正確に解説した図です。
染色体は,原則として同形同大の染色体が2本で1対となり,細胞内で形成・存在します。この同形同大の染色体同士を相同染色体と呼んでいます。さて,減数分裂に先立って全ての染色体は複製され,結果として4本の相同染色体が一時的に存在することになります。この4本の染色体は整然と並んで接合し,二価染色体と呼ばれる構造を作り上げます。第一分裂によって二価染色体は中央から分裂します。さらに第二分裂によって相同染色体同士が分裂し,最終的には染色体数が半減した生殖細胞が4つできあがるという仕組みです(メスの場合,3つは退化します)。
ここで大切なことは,減数分裂は相同染色体間で整然と行われており,他の染色体とは独立な関係であるという事実です。1つの生殖細胞に相同染色体が複数紛れ込むというようなことは起こりません。そこから次のような考察をすることができます。すなわち,2つの色変わり遺伝子に注目し,それぞれの色変わり遺伝子が異なる染色体上に存在しているならば,それぞれの色変わり遺伝子の動きは独立していることになります。逆に,もしも2つの色変わり遺伝子が同一の染色体上に存在しているならば,(特別な変化が起きない限り)2つの色変わり遺伝子の動きは連動していることになります。前者の場合,それぞれの遺伝は互いに相手と何ら関係がなく,独立していると表現されます(1)と2)はこのケースです)。これに対して後者の場合は,片方の遺伝子の動きはもう一方の遺伝子の動きと連動しており,連鎖の関係にあるといいます(3)にはこのケースが含まれます)。3-3 常染色体上の遺伝と常染色体上の遺伝の組合せ
例えば,ホワイトフェイスパイドは常染色体上に存在している色変わり遺伝子を2セット持っていることになります(ホワイトフェイス遺伝子を1セットとパイド遺伝子を1セット)。したがって,ホワイトフェイスパイドをブリーディングに利用しようとするなら,2つの色変わり遺伝子の動きに注目していかなければならないことがわかります。
ここで,大切なことを思い出してください。それは何かというと,遺伝の舞台はあくまでも染色体上なのだという事実です。つまり,2つの色変わり遺伝子に注目して遺伝を考察する場合にも,必ず染色体の動きを追いながら遺伝子の動きを観察しなければならないのです。
具体的に説明を加えていく方が理解しやすいと思いますので,ホワイトフェイスパイドとパイドのペアからどのような雛が生まれるのかについて考えてみることにしましょう(いずれも他の色変わり遺伝子は持っていないと仮定します)。
まず,ホワイトフェイスもパイドも,ともに常染色体上の劣性遺伝であることを押さえます。常染色体上の遺伝ですから雄と雌の区別は重要ではなく,また劣性遺伝ですから2つの色変わり遺伝子がそろって初めて外見上の変化が現れることになります。さらに,常染色体上の色変わり遺伝子は全て別々の染色体上に存在していることも思い出しておきましょう。このようなことから,このペアの遺伝情報を図に示すと,図3-2のようになるわけです。
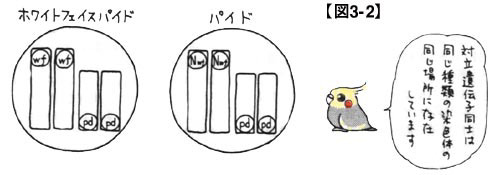
学術的に正確というわけではありませんが,仮に長い方の染色体上にホワイトフェイス遺伝子が乗っているとし,短い方にパイド遺伝子が乗っていると考えておきます。ホワイトフェイス遺伝子とパイド遺伝子は両方とも劣性遺伝子ですから,ホワイトフェイスパイドは,ホワイトフェイス遺伝子を2つにパイド遺伝子2つを,それぞれ別々の常染色体上に持っていることが示されています。同様に,パイドは,(ホワイトフェイス遺伝子の対立遺伝子としての)ノーマル遺伝子を2つにパイド遺伝子2つを,それぞれ別々の常染色体上に持っていると理解できます。
それでは,減数分裂の仕組みをもとに,ホワイトフェイスパイドからどのような生殖細胞ができるのかを考えてみましょう。まず,ホワイトフェイス遺伝子が乗っている長い方の染色体に注目すると,減数分裂の結果,4つバラバラになった相同染色体上のいずれにも,必ずwfが含まれていることがわかります。同様に,パイド遺伝子が乗っていた染色体についても,バラバラになった4つの相同染色体上には必ずpdが存在しています。先にも解説したように,相同染色体以外の染色体は全て独立して分裂しますから,単純にこの2つの結果を組み合わせると生殖細胞の遺伝情報が表現できます。つまり,ホワイトフェイスパイドの生殖細胞には,必ずwf/pdの遺伝情報が伝えられていることになるのです。
パイドについても同じように考えてみると良いでしょう。必ずN(wf)/pdの遺伝情報が生殖細胞に伝えられますね。
この両方の生殖細胞が合体するわけですから,雛の遺伝情報はN(wf)wf/pdpdとなります。つまり,スプリットホワイトフェイスパイドとなり,外見上はパイドとなります。
このように,オカメインコに関する常染色体上の色変わり遺伝では連鎖が一切ないため,常染色体上の2つ以上の色変わり遺伝子に注目しても,それほど複雑になるようなことはありません。
このパターンをしっかりとマスターするために例題1を用意しました。各自で解いてみてください。【例題1】
スプリットホワイトフェイスパイド同士のペアでは,どのような雛がどのような確率で生まれてくるでしょうか。用意したマトリックスを利用して考えてください。
オスの体細胞 N(wf)wf/pdpd ( ) ( ) メスの体細胞
N(wf)wf/pdpd( ) ( ) ( ) ( ) ( ) ( ) 3-4 常染色体上の遺伝と性染色体上の遺伝の組合せ
例えば次のようなペアを考えてみましょう。シナモンパイドのオスとパイドのメスのペアです(他の色変わり遺伝子は持っていないと仮定します)。
まず,パイドは常染色体上の劣性遺伝で,シナモンは性染色体上の劣性遺伝であることに気づくでしょう。続いて,オスは性染色体がZZの組合せ,メスがZWの組合せであることを思い出してください。それぞれ他の色変わり遺伝子は持っていないと仮定していますので,以下のように遺伝情報を表記することが可能です。なお,性染色体上の遺伝子を表記する場合には,存在している色変わり遺伝子のみを書き記せば良かったこと,そして上付で何も書かれていない場合はノーマル遺伝子の存在を意味していることを,それぞれ思い出してください。シナモンパイドのオス;pdpdZ(cn)Z(cn)
パイドのメス ;pdpdZWさて,pdは常染色体上に,cnはZ染色体上に存在していますから,それぞれの遺伝子は独立に動きます。ですから,pdが乗っている染色体の動きとcnが乗っているZ染色体の動きを別々に考察し,それを組み合わせれば精子と卵子の遺伝情報が把握できますね。つまり,オスの精子は必ずpdZ(cn)に,メスの卵子はpdZまたはpdWになることがわかります。
これをマトリックスで示すことにしましょう。図3-3をご覧ください。【図3-3】
オスの体細胞
pdpdZ(cn)Z(cn)pdZ(cn) メスの体細胞
pdpdZWpdZ pdpdZZ(cn) pdW pdpdZ(cn)W pdpdZZ(cn)はスプリットシナモンパイドのオス(外見上はパイド),pdpdZ(cn)Wはシナモンパイドのメスとなります。したがって,この場合も羽色によって雌雄判別が可能となるケースといえます。
第3節との違いは,常染色体の一つを性染色体に置き換えただけです。ですから,基本的な考え方は全く同一で,ただ単に性染色体にまつわる特別な条件を加味しさえすれば,第3節は第4節に変身するわけです。
このケースの例題も用意しました。例題2では生まれてきた雛の羽色から親鳥の遺伝情報を推察しようとしています。今までとはちょっと異なる例題ですが,チャレンジしてみてください。【例題2】
ホワイトフェイス同士のペアリングの結果,まれにホワイトフェイスパールが生まれています。この場合,両親の遺伝情報はどのようなものであると推察されるでしょうか。また,生まれてきたホワイトフェイスパールの雌雄は判別できるでしょうか。3-5 性染色体上の遺伝と性染色体上の遺伝の組合せ
オカメインコの場合,常染色体に関わる色変わり遺伝子は全て異なる染色体上に存在していることが知られています。したがって,2つの色変わり遺伝子に注目する限り,「常染色体上の遺伝と常染色体上の遺伝の組合せ」と「常染色体上の遺伝と性染色体上の遺伝の組合せ」では,それぞれの遺伝子は必ず独立な存在となります。
しかしながら,性染色体は1対しかありませんから,性染色体上で2つの色変わり遺伝子に注目すると,両方の遺伝子は性染色体に乗って同じような動きをするケースが出てきます。つまり,連鎖を考慮しなければならないのです。
以下では,性染色体上の劣性遺伝であるシナモンとパールに注目して,いくつかのパターンを想定してお話を進めていくことにしましょう。3-6 シナモンのオスとパールのメスのペア
便宜的に他の色変わり遺伝子を持っていないものとして議論していきます(以後,同様の仮定をおいておきます)。シナモンのオスは,もはや解説するまでもなく,Z(cn)Z(cn)です。また,パールのメスはZ(pl)Wですね。これを掛け合わせるとどうなるでしょうか。この場合には色変わり遺伝子同士の連鎖を考える必要はないことがわかるでしょうか?
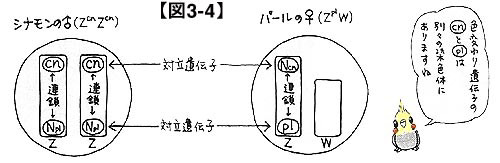
図3-4をご覧ください。染色体と遺伝子を示したおなじみの図ですが,この図をもとに染色体の動きを把握してみてください。cnとplは確かに性染色体に関係していますが,同じ染色体上に存在しているわけではありません。つまり,このような場合はcnとplは独立なのです。したがって,マトリックスは図3-5のようになります。
【図3-5】
オスの体細胞 Z(cn)Z(cn) Z(cn) メスの体細胞
Z(pl)WZ(pl) Z(cn)Z(pl) W Z(cn)W これまでの解説と全く同様の考え方ですね。Z(cn)Z(pl)はスプリットシナモンスプリットパールのオス,Z(cn)Wはシナモンのメスとなります。
ただし,連鎖が全く関係ないかといえば,実はそうでもないんです。図3-4をもう一度ご覧ください。cnとN(pl)は同じZ染色体上に乗っていますね。つまり,cnとN(pl)は連鎖しているのです。同じように,N(cn)とplも連鎖の関係にあります。3-7 シナモンパールのオスとパールのメスのペア
シナモンパールのオスはどのように記号化することができるでしょうか。シナモンもパールも劣性遺伝子ですから,外見上に現れるためには2つ揃っていなければなりません。つまり,図3-6のような状態であるといえます。図3-6に示されているように,シナモンとパールは同一のZ染色体上に存在しているわけです。これを記号化すると,Z(cn/pl)Z(cn/pl)とでも書くことができるでしょう。片方のZ染色体にcnとplが同時に存在しており,もう片方のZ染色体も全く同様です。
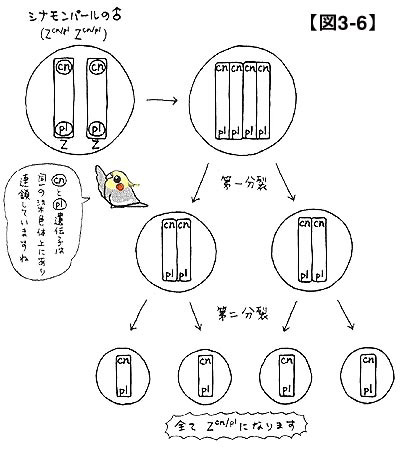
さて,次に減数分裂を考えてみます。染色体の動きに注意しながら精子の遺伝情報を把握してみてください。Z染色体はまず二価染色体を作り,第一分裂と第二分裂を経てバラバラとなり,精子に必ず1本ずつ配分されます。任意のcnに注目すると,そのcnが乗っているZ染色体上に同時に存在しているplと連鎖していることがわかります。Z染色体上に同居しているcnとplは,(特別な変化が生じない限り)同一の精子に配分されるのです。つまり,同じ乗り物(特定のZ染色体)に乗っているのですから,目的地(精子)も同じになると理解してください(ただし,第8節で解説する遺伝子組換えが起こったケースを除く)。
したがって,記号で整理し直すと,Z(cn/pl)Z(cn/pl)からは必ずZ(cn/pl)の精子が作られることになります。Z(cn)やZ(pl),あるいはZが作られることはありませんので,注意が必要です。
パールのメスは今まで通りですので,これらの情報からマトリックスを作成すると図3-7になります。【図3-7】
オスの体細胞 Z(cn/pl)Z(cn/pl) Z(cn/pl) メスの体細胞
Z(pl)WZ(pl) Z(pl)Z(cn/pl) W Z(cn/pl)W Z(pl)Z(cn/pl)はスプリットシナモンパールのオスとなります。plは2つ揃っているのでパールが現れますが,cnは1つしかなく,N(cn)に効果が消されてしまうためです。Z(cn/pl)Wはシナモンパールのメスとなります。
以上のように,色変わり遺伝子が連鎖していても,染色体単位で動きを追っていけば迷うことはありません。混乱したら基本に返るのが遺伝を理解する鉄則なんです。3-8 色変わり遺伝子の連鎖はどうして起こったのだろうか?
第7節で解説したように,遺伝子が連鎖している場合であっても,染色体単位で遺伝子の動きを把握しさえすれば,これまで解説を続けてきた遺伝の基本的な考え方で対処できます。しかし,シナモンとパールがどのようなメカニズムによって同一のZ染色体上に乗るようになったかという点を説明することはできません。シナモンとパールは,同時にまた同地域で発見されたものではありませんから,やはり何らかのメカニズムによって別々に存在していたものが同一のZ染色体に乗るようになったとしか考えられないのです。
図3-1をもう一度ご覧ください。二価染色体が形成されていますが,実はこの二価染色体に秘密があるのです。
二価染色体は減数分裂(第一分裂と第二分裂)によって染色体数が半減します。通常は,正確に相同染色体が二分されるのですが,時として正確な分裂に失敗することがあります。図3-8を見てください。二価染色体の真ん中の2本が絡み合ってしまっていますね。このような状況を乗換えが起こっている,あるいは交叉(こうさ)しているといいます。乗換えが起こったまま第一分裂に入ると,染色体が交わっている点から先でそっくり染色体が入れ替わってしまうのです。つまり,相同染色体間で部分的に情報交換が行われることとなります。これを遺伝子の組換えと呼んでいます。
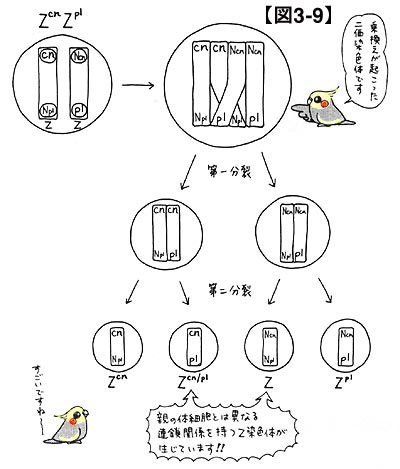
遺伝子の組換えが起こると,当該染色体上の連鎖の関係が変わってしまいます。例えば,Z(cn)Z(pl)の体細胞からは通常Z(cn)かZ(pl)の生殖細胞しか生じないのに,ZやZ(cn/pl)が生じたりもするのです。図3-9を見ていただければ,これらの点が理解できるでしょう。
最近では,人為的な遺伝子組換えによって新しい品種の野菜などが作られていますが,そこまで大袈裟なものではないにせよ,自然界にも遺伝子組換えのメカニズムが存在しているということは驚くべき事実だと感じます。いってみれば分裂の失敗なのですが,それは遺伝子組換えという意義を持ち,同一種の中でも多様な個体を生み出すことにもつながっています。このようなメカニズムが備わっていることによって,環境の変化にも耐えることができる個体の出現を促し,種の保存を巧みに成功させてきたのでしょう。神秘的としかいいようがありませんね。
3-9 遺伝子組換えを考慮したブリーディング
さあ,最も複雑なケースともいえる遺伝子組換えを含んだブリーディングを考察してみましょう。例えば,第6節において行ったブリーディングで得られたオスの雛(Z(cn)Z(pl))とノーマルのメス(ZW)のペアからシナモンパールがとれる確率を考えてみましょう。交叉には,実際には二重・三重と複雑な交叉もありますが,それらは便宜的に無視しておきます。また,10個の生殖細胞(配偶子)あたり,cnとplに関する遺伝子組換えが起こっているものは2個あると仮定します。
以上の条件下では,ZcnZplから作られる10個の生殖細胞の内訳は次のようになります。
Z(cn) ; 4個 Z(pl) ; 4個 Z(cn/pl) ; 1個 Z ; 1個 Z(cn/pl)が作られるということは,遺伝子組換えが起こり,plとN(pl)が交換されたということですから,Z(cn/pl)と同数のZが生じていることになります。つまり,Z(cn)とZ(pl)はそれぞれ40%の確率で生じ,Z(cn/pl)とZはそれぞれ10%の確率で生じる計算となります。
一方,ZWからは,ZとWがそれぞれ50%の確率で生じます。
これらのことから,係数を加えたマトリクスを書いてみます。図3-10です。【図3-10】
|
|
|||||
|
|
|
|
|
||
|
ZW |
|
|
|
|
|
|
|
|
|
|
|
|
この結果,Z(cn/pl)Wだけがシナモンパールのメスとなり,その確率はなんと5%にしか過ぎないことがわかります。10個の精子のうちZ(cn/pl)が1個生じるという高確率を仮定してもなお,結果的にシナモンパールが生まれるのは5%になってしまうわけですから,遺伝子組換えを期待してシナモンパールを作出するのは大変な困難を伴うといえそうです。
3-10 おわりに
これで遺伝の解説は終わりとなります。今回の解説では,注目する色変わり遺伝子を2つにまで拡張しましたが,3つの遺伝子に注目しても,4つの遺伝子に注目しても,考え方は変わりません。つまり,ここまで読み進められた方は,オカメインコの遺伝形式の全てをマスターしたことになるわけです。
オカメインコの品種は今も増え続けているようです。最新の品種の遺伝形式も徐々にわかりつつあります。これらにつきましては情報収集につとめ,またいずれみなさんに報告したいと思っています。
筆者は遺伝の専門家でもなく,解説中には誤りがあるかもしれません。もしも誤りに気づかれましたら,JCCまでご連絡していただけると幸いです。
最後になりましたが,拙劣な文章・解説を我慢して読み続けていただいた方に,心から感謝申し上げます。【主要参考文献】
オカメインコの遺伝特性については,Peggy Cross and Diana Andersen, A Guide to... Cockatiels and Their Mutations ; Their Management, Care and Breeding, Australian Birdkeeper, 1994 を参照いたしました。また,今井逸郎氏がホームページ(http://www.asahi-net.or.jp/‾eq6i-imi/)で公開なさっているオカメインコの遺伝に関する文献から様々な知識を得ています。
遺伝の仕組みについては,メンデルの遺伝に関する一般的なテキストとともに,旺文社 生物辞典[改訂新版]および,岩波 生物学事典 第4版 を利用いたしました。
この他,進化に関するいくつかの著書から貴重な見識を得ています。
【例題1の解答】
オスの体細胞 N(wf)wf/pdpd N(wf)pd wfpd メスの体細胞
N(wf)wf/pdpdN(wf)pd N(wf)N(wf)pdpd N(wf)wfpdpd wfpd N(wf)wfpdpd wfwfpdpd この例題も決して難しくはないですね。ホワイトフェイス遺伝子が乗っている常染色体とパイド遺伝子が乗っている常染色体を別々に考え,その結果を組み合わせれば生殖細胞の遺伝情報は容易に把握されます。それを今度はマトリックスで合体させれば解答が導き出されます。外見上でいいますと,パイドが75%の確率で,ホワイトフェイスパイドが25%の確率で生まれることになります。
両親が外見上でパイドとわかっていますので,その子供に必ずパイドが入ることは直感的にも理解できます。ですから,75%と25%という数字は,スプリットホワイトフェイス同士のペアでホワイトフェイスが出現するかどうかの確率を示しているとも考えることができます。【例題2の解答】
両親がともにホワイトフェイスを示しているので,ホワイトフェイスに関する遺伝情報は容易に把握可能です。問題はパール遺伝子がどうなっているのかという点に絞られます(他の色変わり遺伝子は無視します)。
まず,メスを考えてみましょう。メスの場合,伴性遺伝に関わる色変わりのスプリットは存在していませんから,wfwfZWであることが判明します。どうしてだかわかるでしょうか。もしもwfwfZ(pl)Wなら,このメスはホワイトフェイスパールになってしまいますから,wfwfZW以外は考えらないのです。
そうすると,オスはどうでしょうか。外見上オスはホワイトフェイスですから,パール遺伝子を持っていても1つに限られなければなりません(つまり,スプリットでなければなりません)。また,子供にパールが出ていますから,両親のうち最低限どちらかにパール遺伝子が存在していなければつじつまが合いません。したがって,wfwfZZ(pl)であることが理解できます。
そこでマトリックスを作ってみます。
オスの体細胞 wfwfZZ(pl) wfZ wfZ(pl) メスの体細胞
wfwfZWwfZ wfwfZZ wfwfZZ(pl) wfW wfwfZW wfwfZ(pl)W
唯一,wfwfZ(pl)Wはホワイトフェイスパールとなりますね。その確率は25%です。また,この個体はメスであることも判別できます。
この考察を一歩進めると,外見上ノーマル同士の交配でもホワイトフェイスパールが生まれる可能性があることもわかります。次のようなケースです。
|
|
|||||
|
|
|
|
|
||
|
N(wf)wfZW |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
この結果を見ると,ホワイトフェイスやパールはもちろんのこと,なんとホワイトフェイスパールまで生まれる可能性があるわけです。wfwfZ(pl)Wはホワイトフェイスパールのメスとなります。その可能性は(1÷16×100)で,実に6.25%にしか過ぎません。この可能性は,もちろん上記のような遺伝情報を持った親鳥が前提となっていますから,遺伝情報が管理されていないノーマル同士のペアリングでは,限りなく0%に近いといえそうです。